DNA Replication Stability and Accuracy: The Optimal Engineering Chosen by Cells
Campbell Biology 16.2 Analysis: From the Structural Limitations of the Three Alternative Models to Cellular Traffic Control and Advanced Molecular Biotech

In our previous article, we explored the historical milestones that led to the identification of DNA as the genetic material. Once its identity was established, the scientific community immediately faced the next fundamental question: "How does this simple-looking double helix produce flawless copies with virtually zero error?"
Campbell Biology Chapter 16.2 is far more than a list of replication enzymes to memorize. It reveals a highly coordinated molecular system where the cell balances two conflicting challenges: absolute accuracy and rapid speed. Let us dive deep into the precise biochemical mechanisms behind this process.
1. Fundamental Concepts of DNA Replication: Complementary Base Pairing and Strand Separation
DNA replication relies entirely on the structural features discovered by James Watson and Francis Crick.
Complementary Hydrogen Bonding: The bases inside the DNA double helix pair via specific hydrogen bonds. Adenine (A) always pairs with Thymine (T), and Guanine (G) always pairs with Cytosine (C), ensuring strict complementarity.
Strand Separation: Replication begins when these hydrogen bonds unzip, separating the two strands. Each separated parental strand then serves as a physical template for synthesis.
New Backbone Formation: Free nucleotides align along the exposed parental bases via complementary pairing. They are then covalently linked into a continuous line, forming a stable sugar-phosphate backbone.
2. Three Alternative Models: Why Semiconservative Replication Was Selected
While scientists agreed that genetic information is copied via complementary pairing, they clashed over three competing hypotheses regarding how the physical strands separate and recombine: Conservative, Semiconservative, and Dispersive models.
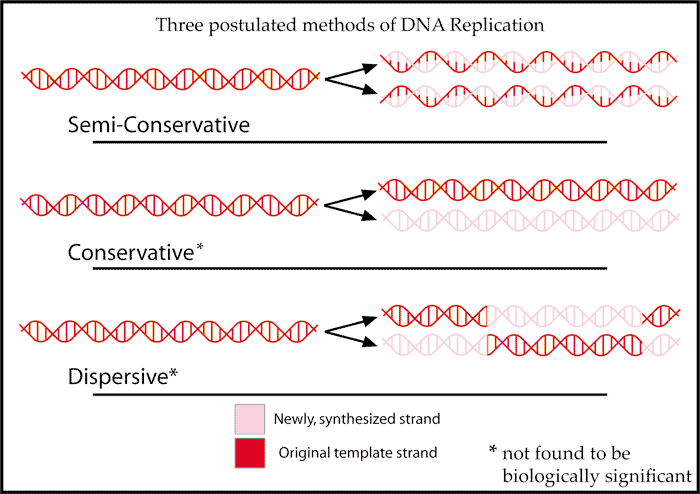
[Image Source: Wikipedia - Semiconservative Model]
(Conservative: Original intact + entirely new double helix / Semiconservative: Half original + half new / Dispersive: Mosaic mixture of old and new pieces)
All three models yield the exact same sequence of genetic data. However, nature strictly utilizes the semiconservative model because it offers superior physical stability and accuracy during the biochemical assembly process.
The Structural Flaw of the Conservative Model
The conservative model posits that the parental double helix remains completely closed—or opens only briefly before snapping back together—while an entirely new double helix is assembled externally.
However, because the template base sequence is sequestered deep inside the tightly wound helix, it is physically impossible for the cell to align and assemble free nucleotides from the outside without direct exposure. Attempting to replicate information without opening the template inevitably causes catastrophic error rates.
The Structural Integrity of the Semiconservative Model
The semiconservative model completely separates the parental strands, exposing the internal bases. Incoming nucleotides bind directly to the exposed template strand via hydrogen bonds before their sugar-phosphate backbones are linked. Because new nucleotides are physically stabilized by the template strand during assembly, the inherent error rate drops drastically.
Furthermore, this structural arrangement allows for real-time error correction (Proofreading). If an incorrect, mismatched base pairs up (such as A with G), the improper hydrogen bonding distance causes a distinct physical distortion or wobble in the double helix. Because DNA Polymerase synthesizes the new strand while it is physically bound to the parental template, it immediately detects this structural distortion, halts progression, and excises the incorrect nucleotide. In a conservative model where assembly happens independently of the original template, this real-time mechanical inspection is structurally impossible.
The Physical Risk of the Dispersive Model
The dispersive model requires cutting the parental DNA into thousands of tiny fragments, synthesizing new segments, and alternatingly stitching them back together.
Repeatedly breaking and reforming covalent phosphodiester bonds introduces a massive risk of double-strand breaks and permanent sequence loss. In contrast, the semiconservative model never cleaves the robust sugar-phosphate backbone of the parental strand; it only unzips the weaker hydrogen bonds between bases, perfectly maintaining the structural continuity of the original DNA.
3. The Meselson-Stahl Experiment (1958)
To distinguish between these three models, Matthew Meselson and Franklin Stahl designed a landmark experiment tracking the physical weight of the DNA strands using heavy nitrogen (15N) and light nitrogen (14N) isotopes.
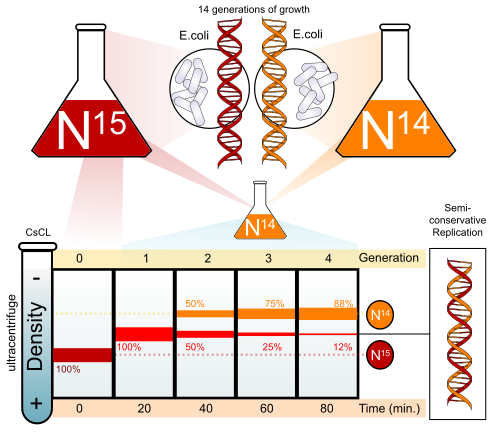
[Image Source: Wikipedia - Meselson-Stahl Experiment]
E. coli cultured in a heavy 15N medium were transferred to a light 14N medium to undergo replication. The density of the DNA was then measured using an ultracentrifuge.
After 1st Generation of Replication: A single "intermediate density" band appeared exactly in the middle of the centrifuge tube. This eliminated the conservative model, which required distinct heavy parental and light daughter bands.
After 2nd Generation of Replication: Two distinct bands appeared: one intermediate band and one completely light band. If the dispersive model were correct, the DNA would have remained a hybrid mixture, producing a single shifting hybrid band indefinitely.
This elegant experiment proved definitively that DNA replicates semiconservatively.
4. Origin of Replication: Speed and Mechanics in E. coli vs. Eukaryotes
The specific site where replication begins is the replication origin. E. coli (bacteria) possesses a single origin of replication, whereas eukaryotic cells (like human cells) utilize thousands. This divergence is driven by genome size, replication speed, and structural physics.
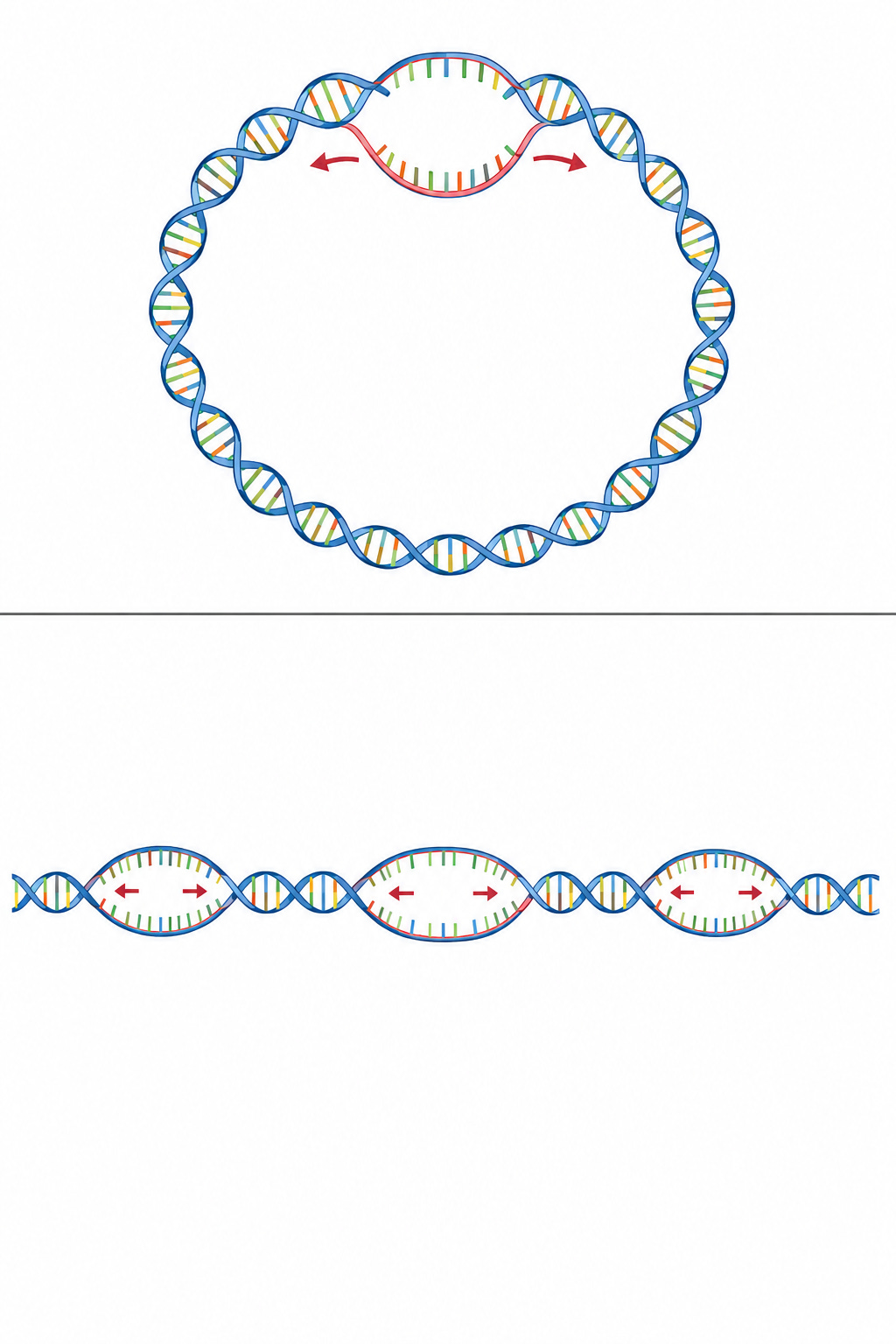
Why E. coli Utilizes a Single Origin
Initiating replication at an origin requires a significant metabolic investment to assemble the necessary initiation proteins and two bidirectional replication forks.
E. coli contains roughly 4.6 million base pairs (bp), which is nearly 700 times smaller than the human genome (3 billion bp). However, bacterial DNA polymerase operates at a blazing speed of roughly 1,000 nucleotides per second. Running just two replication forks from a single origin allows E. coli to fully replicate its entire genome in only 40 minutes, making a single origin highly efficient.
Additionally, E. coli possesses circular DNA. If dozens of replication forks operated simultaneously across a closed circle, the resulting supercoiling and torsional strain would cleave the DNA. Multiple origins would also cause complex physical catenation (interlocking DNA rings) that the cell could not separate during division, resulting in cell death.
Why Eukaryotes Require Thousands of Origins
Eukaryotic DNA polymerases synthesize at a much slower rate of roughly 50 nucleotides per second due to highly rigorous proofreading mechanisms. If a human cell attempted to replicate its 3 billion base pairs using a single origin, the process would take over 800 days, causing the cell to age or die before dividing. To complete replication within the required 8-hour window (S phase), linear eukaryotic chromosomes use thousands of origins to initiate synthesis simultaneously.
Preventing Replication Fork Collisions
Operating multiple replication forks introduces the risk of molecular collisions. When two replication forks moving toward each other meet, they do not crash or damage the template; instead, the two newly synthesized strands are ligated smoothly (fork merger), and the replication machinery disassembles naturally.
A more severe hazard occurs when a fast-moving DNA replication fork collides with an RNA polymerase moving along the same strand to transcribe a gene. To prevent head-on crashes, cells hardwire highly transcribed genes to run in the same direction as the replication fork. If an unavoidable collision occurs and stalls the fork, specialized DNA repair enzymes are recruited immediately to stabilize the structure and restart replication.
5. Summary of Bacterial DNA Replication
Bacterial DNA replication is a highly coordinated multi-enzyme process. Understanding the sequential order of these enzymes reveals the complete blueprint of replication:
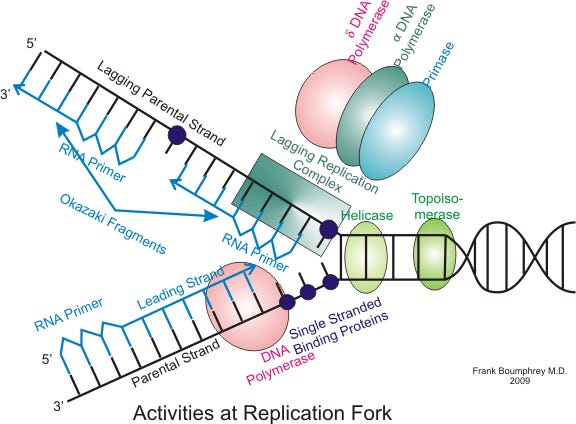
[Image Source: Wikipedia - Eukaryotic DNA replication]
1. Topoisomerase: Relieves the severe overwinding and torsional strain ahead of the replication fork by temporarily cleaving and rejoining the DNA backbone.
2. Helicase: Unzips the double helix by physically breaking the hydrogen bonds between complementary bases.
3. Single-Strand Binding Protein (SSB): Binds to the exposed single strands to prevent them from reannealing or being degraded by nucleases.
4. Primase: DNA polymerase cannot initiate synthesis on bare single-stranded DNA; it requires an existing strand to build upon. Primase synthesizes a short RNA primer (approx. 10 nucleotides) to provide the initial starting point.
5. DNA Polymerase III: The primary synthesis enzyme. It uses the 3'-OH end of the RNA primer as an anchor to continuously append complementary DNA nucleotides, building the new strand.
6. DNA Polymerase I: The finishing enzyme. It removes the temporary RNA primer nucleotides and replaces them with corresponding DNA nucleotides.
7. DNA Ligase: Seals the remaining structural nicks between newly synthesized fragments (such as Okazaki fragments on the lagging strand), creating a continuous phosphodiester backbone.
6. Nucleotide Excision Repair
Even after successful replication, environmental factors like ultraviolet (UV) radiation or carcinogens constantly distort the DNA structure. UV light frequently causes adjacent Thymine bases to covalently bind together, forming a "thymine dimer" that creates a structural bulge. The cell deploys a repair pathway to correct this damage before the next replication cycle:
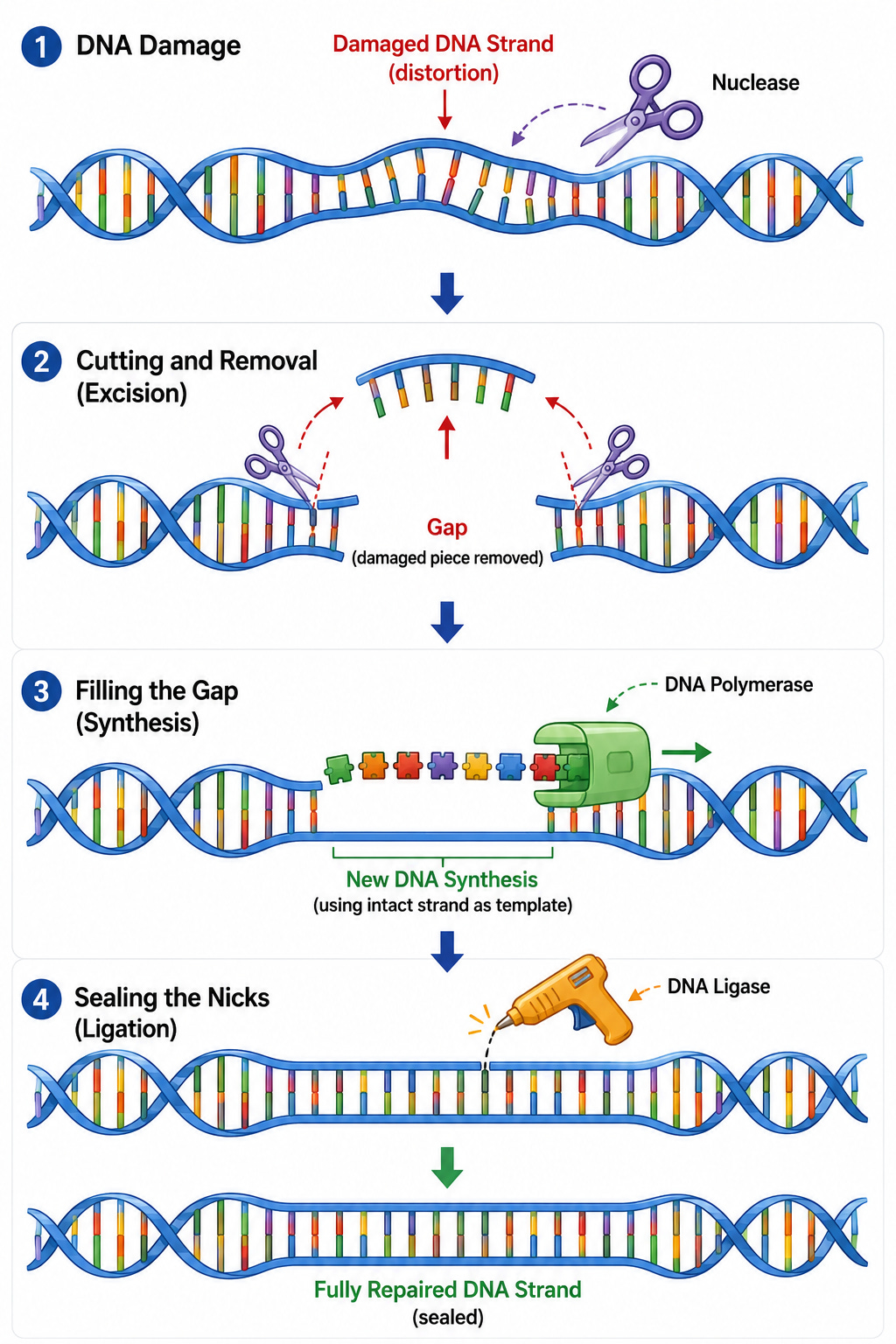
Nuclease: Detects the structural bulge caused by the lesion, cuts the damaged strand on both sides of the error, and removes the affected oligonucleotide fragment entirely.
DNA Polymerase & DNA Ligase: DNA Polymerase uses the undamaged opposing strand as a template to fill in the missing nucleotides, and DNA Ligase seals the backbone to restore the original strand.
7. The End-Replication Problem and Telomeres
Eukaryotic linear chromosomes face an inevitable structural dilemma: the DNA molecule shortens with every round of replication.
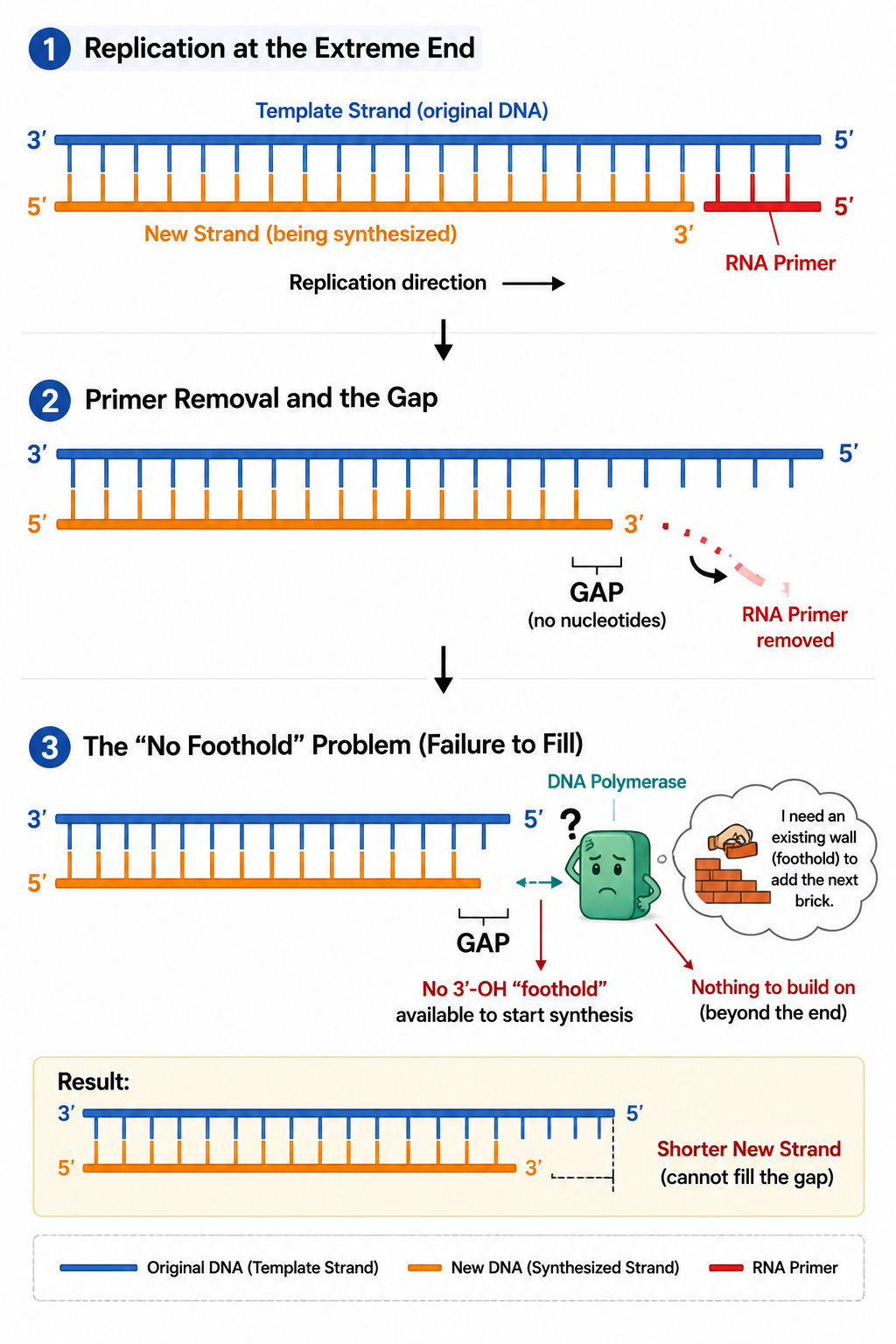
The End-Replication Problem
During linear DNA replication, an RNA primer sits at the absolute 5' end of the new strand to initiate synthesis. Once replication concludes, this RNA primer is removed.
DNA polymerase can only append nucleotides to an existing 3'-OH group. Because this missing primer was at the absolute start of the new strand, there is no upstream 3'-OH group available for a polymerase to bind to and fill the gap. Consequently, the newly synthesized daughter strand is always shorter than the parental template by the length of the primer. If critical genes were located at these extreme ends, essential genetic data would vanish within a few cell divisions.
Telomeres
To shield vital genetic information, eukaryotes feature telomeres: long stretches of non-coding, repetitive nucleotide sequences (TTAGGG in humans) at the terminal ends of chromosomes. This acts as a protective buffer sequence.
The structural region where primers are removed and left unfilled resides entirely within this non-coding telomeric zone. As the chromosome shortens with each replication cycle, only these repetitive, non-functional telomeric sequences are lost, keeping core functional genes completely unharmed.
The Hayflick Limit and Aging
This protective buffer is finite. Normal human somatic cells can divide roughly 50 to 70 times before the telomeres degrade entirely. Reaching this threshold—known as the Hayflick Limit—triggers a cellular alarm. The cell permanently halts division and enters a state of senescence, which represents the molecular basis of biological aging.
Telomerase and Cancer
Sperm, eggs, and somatic stem cells must divide continuously without losing genetic integrity. These specific cells express an enzyme called Telomerase, which actively appends repetitive sequences back onto the shortened telomere ends, maintaining their length indefinitely.
However, malignant cancer cells exploit this exact mechanism. By abnormally mutating to reactivate telomerase, cancer cells bypass the Hayflick Limit entirely, achieving replicative immortality and dividing uncontrollably.
In our next article, we will examine how these exact principles of replication mechanics and fork kinetics are translated into multi-billion dollar platform technologies. We will break down the precise business models and technological competitive advantages of market leaders like PacBio and next-generation oncology biotechs. If you want to see how fundamental molecular engineering drives the future of medicine and industry, stay tuned for the next installment!