Exploring Chromatin packing in a Eukaryotic Chromosome

Genetic information stored in DNA is not simply a linear strand of molecules. To pack roughly 2 meters of DNA into a microscopic nucleus spanning only a few micrometers, the cell utilizes complex protein interactions while simultaneously maintaining precise control over gene expression.
This sophisticated packaging technology begins with overcoming the fundamental physical properties of DNA and extends to dynamic structural transitions in response to the drastic environmental changes of cell division.
1. The Foundation of Chromatin Packaging: The Double Helix and Histone Electrostatic Interactions
The starting point of DNA packaging is the double helix structure. In a ribbon model of DNA, each ribbon represents an individual polynucleotide strand. When completely free of bound proteins, this naked DNA molecule measures approximately 2 nm in diameter.
The primary responsibility for packaging this long, thin molecule falls on histone proteins. Histones possess unique chemical properties: more than 20% of their amino acids consist of positively charged lysine (Lys) and arginine (Arg) residues. Because the phosphate backbone of DNA carries a strong negative charge, a highly stable and tight association is formed between them through powerful electrostatic interactions.
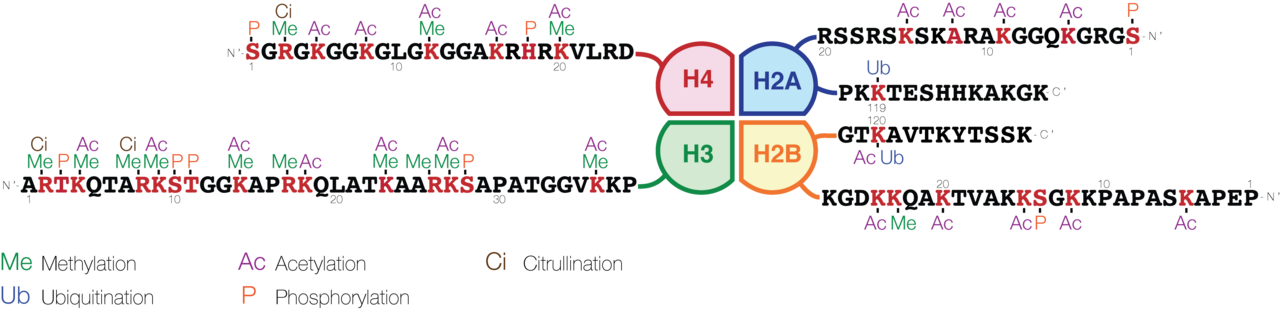
As key players in maintaining genomic structure, histones are highly conserved across eukaryotes, showing very little sequence variation. The most common histones found within chromatin are the four core types: H2A, H2B, H3, and H4. Two of each type associate to form an octameric protein core, which acts as a molecular spool for the DNA double helix to wrap around. Additionally, the linker histone H1 binds to this structure to lock the wrapped DNA in place and facilitate higher-order folding.
[Image Source : Wikipedia - Nucleic acid double helix]
2. Organization of the 10-nm Fiber: The Roles of Nucleosomes and Linker DNA
In its partially unfolded state, chromatin forms a fiber approximately 10 nm in diameter, which appears under an electron microscope as "beads on a string." The basic unit of this genomic packaging, representing each individual "bead," is called a nucleosome.
A single nucleosome consists of the octameric histone core (formed by H2A, H2B, H3, and H4) wrapped nearly twice (specifically 1.67 turns, roughly 146 base pairs) by the DNA double helix. The amino-terminal ends of the core proteins, known as histone tails, protrude outward from the nucleosome. Reversible chemical modifications of these tails—such as methylation and acetylation—serve as key epigenetic switches that regulate gene expression.
Connecting these nucleosome beads is a segment of exposed, double-stranded DNA called linker DNA. Typically ranging from 20 to 80 base pairs in length, this linker region is where the H1 histone binds, acting as a physical trigger to condense the 10-nm fiber into more compact structures like the 30-nm fiber.
Rather than being static, this packaging is highly dynamic. During DNA replication, the nucleosome must temporarily disassemble to allow the replication machinery to unwind and read the double helix.
To prevent the displaced histone proteins from dispersing into the nucleoplasm, they are immediately captured and stabilized by histone chaperones (such as ASF1 and CAF-1) waiting near the replication fork. As soon as the replication machinery passes and the new daughter strand is synthesized, these chaperones promptly reassemble both the parental and newly synthesized histones back onto the DNA. A similar chaperoning process occurs during transcription, where histone displacement ensures that RNA polymerase has safe, temporary access to the DNA template.
[Image Source : Wikipedia - Histone]
3. Genomic Compartmentalization in Interphase: The Boundary Between Euchromatin and Heterochromatin
When the cell is not dividing (during interphase), chromatin is partitioned into two distinct density states based on its transcriptional activity:
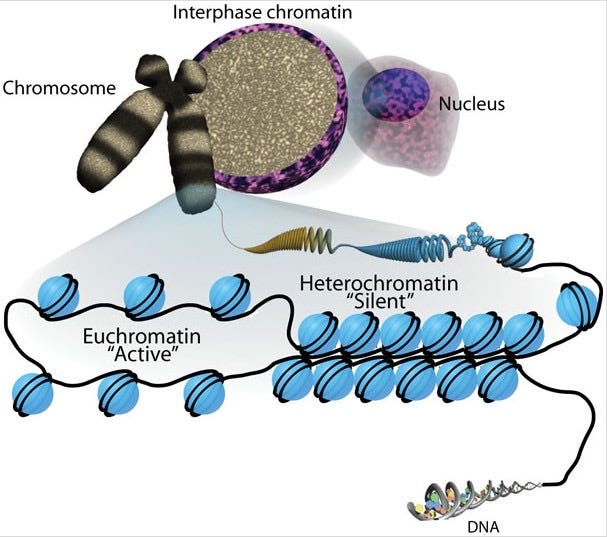
Euchromatin: Regions where the 10-nm fiber is loosely arranged in a more open configuration. Because transcriptional machinery has easy physical access to the DNA, the genes within these regions are actively expressed (though a transient 30-nm fiber organization may occur depending on cell type or timing).
Heterochromatin: Regions where the 10-nm fiber is highly compacted and densely packed. This high density physically blocks transcriptional proteins, meaning genes within heterochromatin are generally silenced (not expressed).
It is a common misconception that heterochromatin simply represents a more "macroscopic" view of euchromatin, but this is not a matter of viewing scale. Under the same magnification, euchromatin and heterochromatin are distinguished by their local physical and chemical density. Within the exact same chromosome (a single continuous DNA molecule), some regions are kept open for active use (euchromatin), while others (such as centromeres and telomeres) are tightly locked down for structural integrity (heterochromatin). Rather than a "zoom in or zoom out" scale difference, it is a functional distribution of open and closed domains organized by the cell.
This precise spatial boundary and 3D organization are established by heterochromatin-binding proteins like HP1 (Heterochromatin Protein 1), along with regulatory enzymes like histone methyltransferases (HMTs) and histone acetyltransferases (HATs). Additionally, architectural proteins like CTCF and cohesin organize the DNA into loops to insulate these active and inactive domains.
Remarkably, this precise compartmentalization disappears the moment mitosis begins. When cell division starts, all transcriptional activity is halted (transcription shutdown). The cell's sole priority shifts from fine-tuning gene expression to condensing and cleanly segregating its duplicated genetic material. Consequently, the epigenetic boundary proteins dissociate from the chromosomes, and the entire genome is subjected to uniform, high-intensity compaction, temporarily dissolving the distinct interphase compartments.
[Image Source : Wikipedia - Euchromatin]
4. Mechanical Compaction During Mitosis: The Dynamic Actions of Condensins I and II
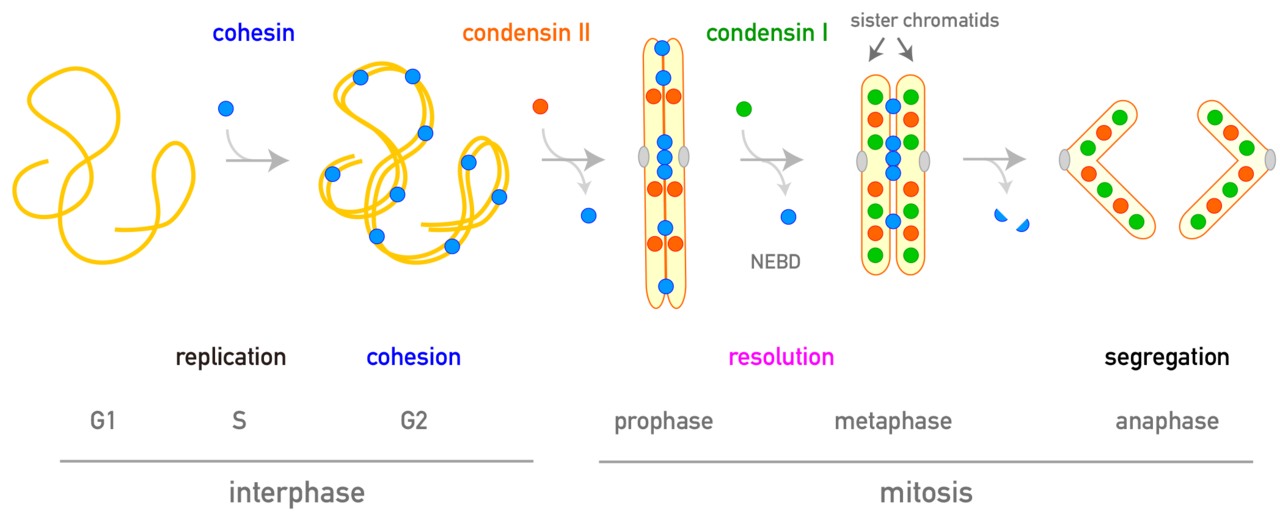
Because DNA replication is completed during S phase, a cell entering mitosis possesses chromosomes that consist of two identical sister chromatids. During prophase and prometaphase, these flexible chromatin fibers undergo extreme physical compaction coordinated by two related multi-protein complexes: condensin II and condensin I.
4-1. Prophase: Condensin II and Primary Loop Formation
At the onset of prophase, the chromatin of each sister chromatid begins to condense. First, condensin II complexes bind to the 10-nm chromatin fiber and initiate the process of loop extrusion, forming increasingly larger DNA loops. These condensin II complexes align longitudinally to construct a central scaffold that runs down the axis of the chromosome, with the extruded loops radiating outward from it.
These loops serve as the structural backbone of chromosome condensation. Simply folding a long DNA strand without order would lead to severe tangling; however, anchor-looping the strand onto a central axis minimizes volume while keeping the DNA organized. As these loops grow outward, the chromosome becomes wider but experiences dramatic axial shortening, reducing its longitudinal length to about half of its interphase state.
4-2. Prometaphase: Condensin I and Helical Twisting of the Axis
Once prometaphase begins and the nuclear envelope breaks down, condensin I complexes in the cytoplasm gain access to the chromosomes. Condensin I does not build a separate scaffold; instead, it binds to the outer regions of the loops already established by the condensin II central scaffold.
Condensin I functions by subdividing the large, pre-existing loops into nested, smaller loops. This can be compared to organizing a suitcase: rather than packing large, bulky items loosely, dividing the space with smaller packing organizers allows much more material to be tightly packed within the same volume, drastically increasing the overall density.
As this nested looping continues, the chromosome becomes shorter, wider, and denser. Simultaneously, the central condensin II scaffold itself begins to twist into a helix. This helical twisting does not elongate the chromosome; rather, it drives it into an extremely compact, rigid state.
Just like a coiled telephone cord occupies a fraction of the length of a stretched-out wire, the twisting of the central axis allows a far greater number of DNA loops to be packed per unit length.
This step-by-step looping and helical twisting serve two crucial evolutionary purposes: mechanical protection and efficient transport. When the spindle apparatus exerts powerful pulling forces on the sister chromatids during metaphase, the chromosomes must possess exceptional tensile strength to prevent double-strand breaks or physical tearing. Compacting the loose chromatin into dense, organized packages also ensures that no DNA is lost or damaged during movement. Rather than a redundant cycle of shortening and lengthening, this is a highly directed, continuous compaction pathway designed to protect the genome as it moves toward its final destination at the metaphase plate.
By metaphase, the chromosome reaches peak compaction with the maximum density of loops per turn, resulting in its shortest and most robust physical state, with the sister chromatids fully resolved for segregation.
[Image Source : Wikipedia - Condensin]
5. Spatial Arrangement in the Interphase Nucleus: Chromosome Territories
Once cell division is complete and the cell returns to interphase, the highly condensed metaphase chromosomes unpack back into loose chromatin fibers within the newly formed nucleus.
Using a molecular visualization technique known as chromosome painting—which labels individual chromosomes with distinct combinations of fluorescent probes—researchers can track the positions of these unpacked fibers. This imaging reveals that even in their loose, thread-like interphase state, chromosomes do not float around in a chaotic, tangled mess.
Instead, each chromosome occupies a distinct, non-overlapping 3D region within the nucleoplasm called a chromosome territory. Interestingly, the two homologous chromosomes of a diploid pair do not pair up; they are typically located in completely different regions of the nucleus.
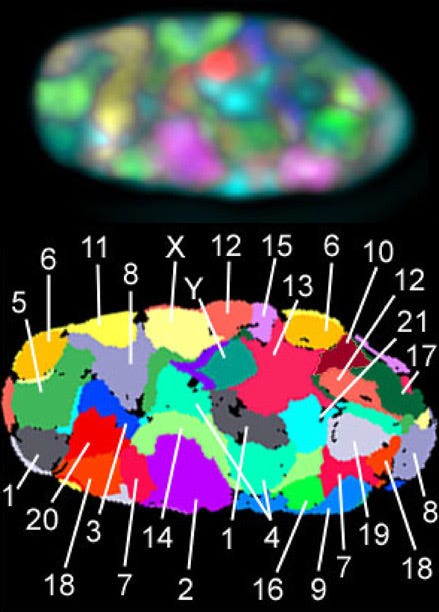
[Image Source : Wikipedia - Chromosome territories]
There are vital biological reasons why chromosomes maintain these strict physical boundaries:
1. Preventing Genomic Damage: If the 46 giant DNA molecules inside a human cell nucleus were completely mixed like a bowl of spaghetti, the replication and transcription machinery would constantly cause physical stress as they unwound the strands. This would lead to catastrophic entanglements, double-strand breaks (DSBs), or abnormal chromosome translocations.
2. Structural Anchoring: The inner nuclear membrane is lined with a supportive protein meshwork called the nuclear lamina, which connects to an internal structural network known as the nuclear matrix. Specific regions of each chromosome are chemically anchored to these structures, keeping them physically tethered in place and preventing them from drifting.
3. Optimizing Gene Regulation: When genes need to be transcribed, only the specific active regions of a chromosome loop outward from their designated territory to interact with shared transcription machinery (known as transcription factories). This organized compartmentalization ensures that gene expression is carried out with maximum speed and energy efficiency.