Unlocking the Secrets of Heredity: the Epic Journey of DNA Discovery
A Deep Dive into Campbell Biology 16.1: The Pivotal Moments that Overturned the Protein Theory and Ushered in the Era of Molecular Biology

Since the dawn of agriculture, humanity has intuitively understood that traits are passed down from parents to offspring. However, until the early 20th century, no one knew exactly what substance inside the cell was responsible for this miraculous phenomenon. At the time, scientists firmly believed that proteins—composed of 20 different kinds of complex and diverse amino acids—were the genetic material. DNA, by contrast, was dismissed as a molecule far too simple to encode vast amounts of hereditary information, as it consists of only four repeating nucleotides.
Today, we will trace the chronological timeline of Campbell Biology 16.1 to explore the dramatic moments that shattered these preconceptions and conclusively proved that DNA is the true code of life.
1. Griffith’s Experiment (1928): The Mysterious Discovery of 'Transformation'
While attempting to develop a vaccine against pneumonia, Frederick Griffith stumbled upon a historic breakthrough. He worked with two strains of Streptococcus pneumoniae:
S (Smooth) Strain: Possesses a smooth, protective polysaccharide capsule that shields it from the host's immune system. It is pathogenic (lethal to mice).
R (Rough) Strain: Lacks this protective capsule, making it non-pathogenic (harmless to mice).
Griffith injected heat-killed S strain bacteria into mice, and as expected, the mice survived. However, when he mixed these heat-killed S cells with live, harmless R strain cells and injected the mixture, the mice unexpectedly died of pneumonia. Even more shockingly, live S strain bacteria were recovered from the blood of the dead mice.

[Image Source: Wikipedia - Griffith’s Experiment]
(Live S strain -> mouse dies / Live R strain -> mouse lives / Heat-killed S strain -> mouse lives / Mixture of heat-killed S and live R strains -> mouse dies and live S strain recovered)
The results were stunning. An "unknown chemical component" from the dead S cells had permanently transformed the living R cells into deadly S cells. Griffith termed this phenomenon transformation.
But wait—why did this genetic material remain perfectly intact even after the S cells were killed by heat?
Proteins, which scientists at the time believed to be the hereditary carrier, are highly vulnerable to heat. High temperatures cause them to denature—losing their intricate three-dimensional structures and, consequently, their biological functions. If proteins were the genetic material, the heat-killed S cells would never have been able to transform the R cells.
DNA, however, is structurally resilient. The rugged backbone made of alternating sugars and phosphates is held together by strong covalent bonds that easily withstand such heat. While the weaker hydrogen bonds between the strands do break under high temperatures, causing the double helix to temporarily unzip, they effortlessly snap back together once the temperature drops. The blueprint remained fully functional. The R cells simply absorbed this floating DNA from the dead S cells, integrated it into their own genome, and began manufacturing the deadly protective capsule according to the stolen blueprints.
2. Hershey and Chase’s Experiment (1952): The Definitive Proof That "DNA is the Genetic Material"
Following Griffith's work, Oswald Avery and his colleagues chemically proved that the transforming principle was DNA, yet a skeptical scientific community remained unconvinced. The final blow to the protein theory came from Martha Chase and Alfred Hershey's legendary experiment using the T2 bacteriophage.
Why was the T2 phage the perfect tool?
This virus has a devastatingly simple anatomy, making it an ideal candidate to isolate the identity of genetic material:
1. Outer Shell: Composed of 100% protein. (Can be selectively tagged with radioactive Sulfur-35)
2. Inner Core: Composed of 100% DNA. (Can be selectively tagged with radioactive Phosphorus-32)
When a T2 phage infects an E. coli bacterium, it injects its genetic material inside while leaving its empty shell stranded on the outside. If the material injected inside the cell turned out to be protein, then protein was the genetic material; if it was DNA, then DNA won the title.
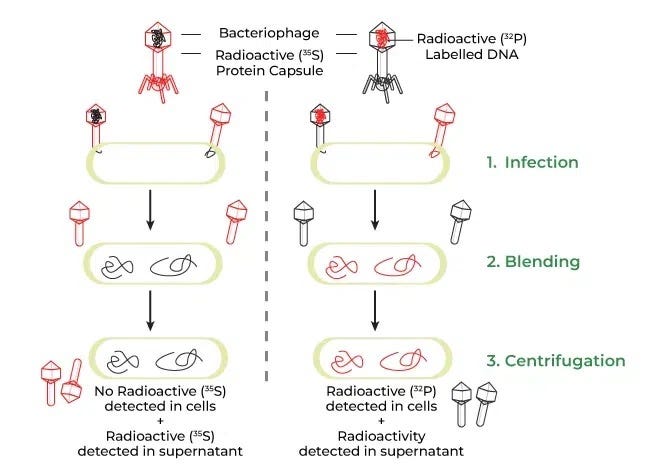
[Image Source: Wikipedia - Hershey and Chase’s Blender Experiment]
(Infection by Sulfur-35-labeled protein phages and Phosphorus-32-labeled DNA phages, agitation in a kitchen blender, centrifugation to separate extracellular phages from bacterial cells, and measurement of radioactivity in the supernatant vs. pellet)
Hershey and Chase separately prepared phages with radioactive labels on either their protein coats or their DNA, then allowed them to infect E. coli. Using a standard kitchen blender, they sheared the empty phage coats off the bacterial surfaces, then spun the mixture in a centrifuge to force the heavy bacterial cells to form a pellet at the bottom.
The results left no room for doubt. The radioactivity from Phosphorus-32—which labeled the DNA—was found strictly inside the bacterial pellet. Conversely, the Sulfur-35 from the protein coats remained in the liquid supernatant. This elegantly demonstrated that DNA was the sole molecular entity injected by the virus to hijack the cell and direct the production of new phages.
3. The Molecular Architecture of DNA: The Biochemical Inevitability of Directionality and Stability
Once DNA was crowned the genetic material, scientists rushed to answer a new question: "How can such a structurally simple molecule store vast amounts of information and replicate itself?" To figure that out, we must examine the DNA strand at a nanoscale level.
The Sugar-Phosphate Backbone: The Stability of Negative Charges
DNA is built by stacking repeating building blocks called nucleotides vertically. Each nucleotide consists of a phosphate group, a five-carbon sugar (pentose), and a nitrogenous base (A, T, G, or C).
Crucially, the phosphate groups carry a net negative charge. This makes DNA highly soluble in the aqueous environment of the cytoplasm and, because like charges repel, prevents the massive strands from tangling awkwardly, keeping the structure uniform and stable.
Horizontal for Information, Vertical for Support: Why DNA Stacks Upwards
As nucleotides chain together, the sugar of one nucleotide fastens tightly to the phosphate of the next via a rugged covalent bond called a phosphodiester linkage. This creates an incredibly rigid vertical backbone capable of enduring chemical and thermal stress. On the flip side, the horizontal links—the nitrogenous bases meeting in the middle—are bound by loose, weak hydrogen bonds. This structural contrast is deliberate: the horizontal bonds must unzip effortlessly whenever the cell needs to read or replicate its genetic data. The vertical axis secures structural integrity, while the horizontal axis handles division and information.
But why must they stack so tightly on top of each other?
The nitrogenous bases (A, T, G, C) are fundamentally hydrophobic (water-fearing). If DNA were a loose, sprawling horizontal ladder, these hydrophobic bases would constantly be exposed to surrounding water molecules, destabilizing the system. By stacking them vertically face-to-face (base stacking), the bases shield each other from water. This tight arrangement triggers weak but massive collective van der Waals attractions between the stacked rings, locking the DNA into a highly compact, warp-free, and rigid column.
Heads and Tails: The Biochemical Mandate Behind 5' -> 3' Replication
A DNA strand has a strict, uncompromising directionality. By numbering the carbons on the sugar ring, we designate the end with the exposed phosphate group attached to the 5th carbon as the 5' end (the head), and the end with the free hydroxyl (-OH) group on the 3rd carbon as the 3' end (the tail).
In every living organism, DNA replication occurs exclusively in the 5' -> 3' direction. This means an incoming nucleotide can only attach its head to the 3'-OH tail of the existing strand. This isn't an arbitrary whim of nature; it's a profound energetic necessity.
What if the factory ran backward?
In standard 5' -> 3' synthesis, the incoming nucleotide brings its own fuel: a high-energy triphosphate (P-P-P) tail on its 5' carbon. If a typo occurs, the proofreading enzyme clips the mismatch out, leaving a clean, standard 3'-OH on the strand. The next incoming nucleotide simply walks up, wielding its own triphosphate pack, and replication resumes smoothly.
In a hypothetical 3' -> 5' synthesis, the fuel tank would have to sit on the existing strand's 5' tip. If the proofreading machinery snipped a mismatched nucleotide out of this backward strand, the act of cleavage would inevitably carry away the high-energy triphosphate tail from the strand's tip. The strand would be left stranded with a dead, low-energy monophosphate (P). When the next incoming nucleotide approaches with its 3'-OH, there is no energy left on the strand to forge a new bond. The replication factory permanently stalls. In short, the vital requirement for proofreading forced evolution to completely abandon the 3' -> 5' pathway.
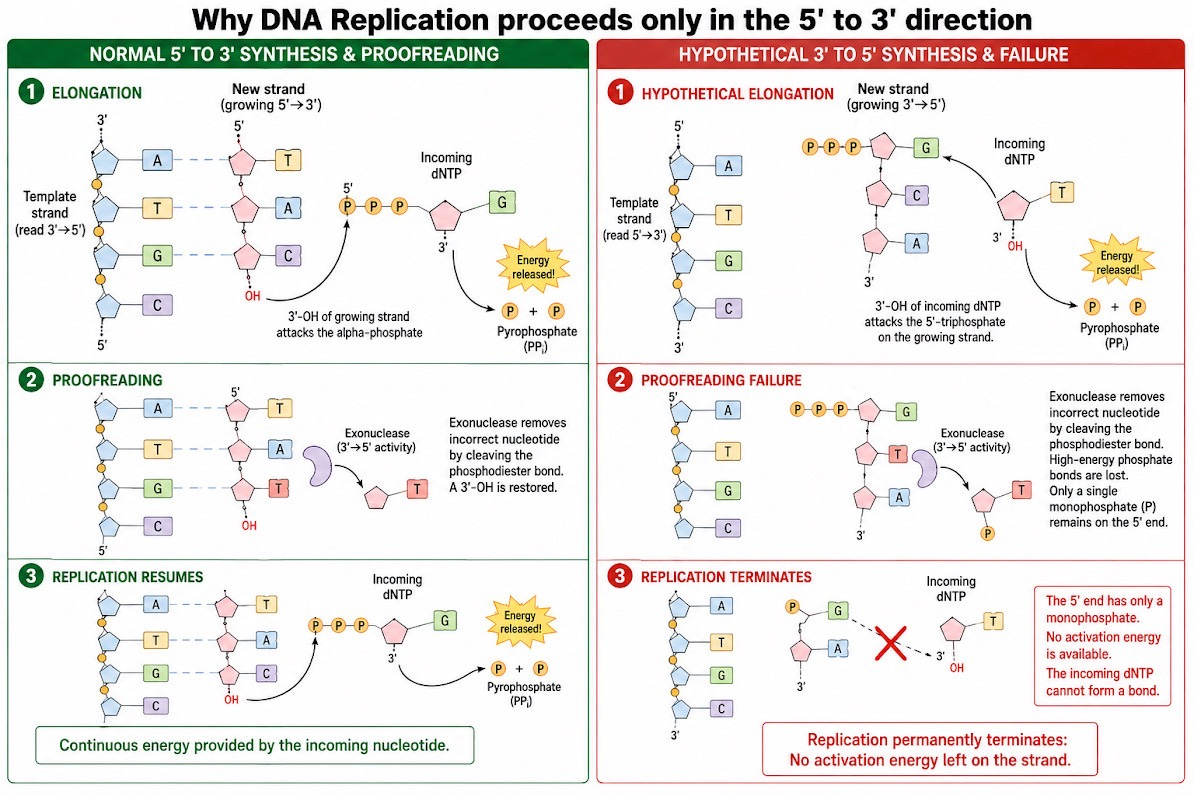
(Visualizing the chemical blocks showing the energy depletion that halts replication in the hypothetical right panel)
4. Rosalind Franklin’s X-Ray Image and the Completion of the Double Helix
By the early 1950s, scientists knew DNA was an elongated chain, but they were blind to its 3D architecture. Was it a single strand? A triple helix? Were the bases facing inside or out? Rosalind Franklin’s historic X-ray diffraction photograph finally pierced through the darkness.
Why the 'X' Pattern Signaled a Helical Structure
Franklin beamed X-rays through crystallized DNA fibers, capturing their scattering pattern on film. The result was a stark, beautifully symmetric 'X' shape. To trained physicists, this 'X' shouted a singular truth: "This molecule is a twisted helix, shaped like a spiral spring!" Why?
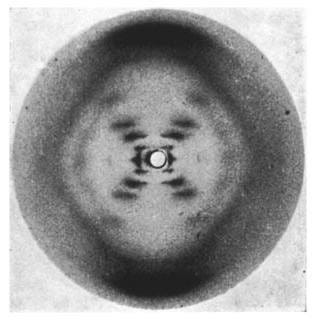
[Image Source: Wikipedia - Photo 51]
Imagine if DNA were a flat, straight, two-dimensional ladder. The sugar-phosphate backbones would stand perfectly vertical, and the flat base rungs would run strictly horizontal. If you shot an X-ray beam through this flat ladder, the light would strike the horizontal rungs perpendicularly. The exiting light would diffract evenly up, down, left, and right, printing parallel horizontal lines or grid-like dots on the film. There would be absolutely no reason for the light to twist into diagonal lines.
Now, imagine a spring coiling upward. Trace the slope of the wire:
1. On the front side: The wire ascends at a diagonal angle rising to the right (forward slash shape).
2. On the back side: As it loops around, the wire climbs at a diagonal angle rising to the left (backslash shape).
When X-rays hit these tilted, repeating paths of the backbone, the light deflects perpendicular to those precise slopes. The right-leaning front strands deflect light diagonally to one side, while the left-leaning back strands deflect it to the opposite side. The overlapping reflections of thousands of coiling strands stack up on the film, creating a stark, intersecting 'X' pattern.
The Rhythmic Dots: Evidence of a Perfect Turn
Looking closer at Photo 51, the 'X' isn't made of smooth brushstrokes; it is composed of dots spaced at strict, rhythmic intervals. This proved that the helix has a perfectly uniform pitch—the vertical distance it takes for the spiral to complete one full turn. Because the atomic intervals repeat with mathematical precision, the scattered X-rays interfered rhythmically to produce cleanly separated dots.
Furthermore, calculating the math behind this diffraction pattern revealed that the diameter of the DNA cylinder was an immutable, constant 2 nanometers wide from top to bottom.
Chargaff’s Ratio and Watson’s Eureka Moment
The final puzzle piece lay in how the bases paired inside that 2-nanometer cylinder. Erwin Chargaff had previously discovered an invariable ratio across all organisms: the amount of Adenine always equals Thymine, and the amount of Guanine always equals Cytosine (A% = T%, G% = C%).
James Watson and Francis Crick originally assumed that bases paired like-with-like (A with A, G with G). However, this created a major structural problem. Adenine and Guanine are large, double-ringed structures (purines), while Thymine and Cytosine are small, single-ringed structures (pyrimidines). Pairing purine-with-purine made the ladder bulge out, while pyrimidine-with-pyrimidine made it pinch inward. This glaringly violated Franklin's proof of a perfectly uniform 2-nanometer diameter.
By combining all these clues, Watson realized that a large purine must always pair with a small pyrimidine to keep the width constant. Guided by Chargaff's ratios, he found that Adenine fits perfectly with Thymine, and Guanine with Cytosine via complementary hydrogen bonds. The double helix was complete.
From Griffith's accidental observation of transforming bacteria to Franklin's masterful X-ray capture and Watson and Crick's brilliant structural synthesis, the story of DNA is one of the most elegant, interconnected dramas in scientific history. The next time you look at a diagram of the double helix, remember that its shape is a testament to the uncompromising chemical laws and survival strategies operating inside our cells.